 |
| Electrodos de marcapasos conectados a un torso (dominio público) |
Este potencial de acción especial es producido por las células que se encuentran en el nodo sinoatrial, también conocidas como células marcapasos.
Las células marcapasos propiamente tales no tienen potencial de reposo, sino que un potencial diastólico máximo. Apenas alcanzan la repolarización, y al llegar a su potencial más negativo (-55 mV para el NSA y -60 mVpara el NAV) inmediatamente se comienza a despolarizar de forma instantánea (potencial marcapasos). Esto ocurre por varios canales, siendo el más importante el de corriente F (IF, o canal funny), que es un canal que permite la entrada de Na+ cuando se alcanza el potencial más negativo, y que están abiertos virtualmente durante todo el potencial marcapasos.
Hay corrientes menos
significativas como la INa-B, que es una corriente menor
de sodio que existe en el reposo de estas células o en el potencial
marcapasos.
En la parte final del potencial
de marcapasos, al acercarse al umbral de los canales Ca2+
L se inicia la apertura de los canales Ca2+ tipo T (de
apertura e inactivación rápidas).
La fase 0 de estas células no
es dada por el Na+, sino que por Ca2+.
Sus diferencias más
significativas son:
- Potencial diastólico máximo.
- Potencial de marcapaso, corriente IF.
- Fase 0: ICaT más precoz, ICaL permite fase 0 y meseta.
- Fase 3: Regularización por canales de K+.
 |
| Potencial de acción de célula marcapasos y sus fases (Diberri, Wikimedia Commons. CC BY-SA 3.0). |
Regulación de la contracción y regulación cardíaca.
El PA ingresa a as células
cardíacas por los túbulos T, llegando a las diadas, donde hay
cercanía con los receptores de dihidropiridina (DHP, canales de Ca2+
tipo L) y receptores de rianodina, donde está el típico
sistema de acoplamiento-contracción cardíaco de liberación de
Ca2’activada por Ca2+. Este Ca2+
difunde por los miofilamentos donde interactúa con la troponina
C, que modifica al complejo troponina desplazando a la
tropomiosina, permitiendo la interacción actina-miosina.
 |
| Esquema que muestra las proteínas claves del ciclo cardíaco del calcio y del acoplamiento excitación-contracción (Wikimedia Commons, CC BY-SA 4.0) |
La troponina C tiene una
afinidad por el calcio que se puede modificar: la fosforilación (por
PKA) mejora la afinidad, que aumenta en presencia de catecolaminas.
La cantidad de Ca2+
que se libera depende de la concentración dentro del retículo, y
este a su vez depende de la bomba de Ca2+ del retículo
(que tiene un regulador negativo, el fosfolambano).
El fosfolambano no fosforilado
inhibe a la bomba de Ca2+ del retículo, pero, al hacerlo,
las catecolaminas dejan de inhibirla y aumenta la velocidad de
reposición del Ca2+, lo que tiene dos efectos
importantes:
- Durante la diástole la recaptación de Ca2’hacia el retículo es rápida, permitiendo una mejor relajación.
- Durante la sístole, como aumenta la concentración de ión calcio dentro del retículo, mejora la disponibilidad de ión calcio libre y la contractibilidad.
Las catecolaminas:
- mejoran la sensibilidad al Ca2+.
- Mejoran la captación de Ca2+ → la relajación hacia el retículo.
- Mejoran la liberación de ión calcio en el siguiente latido.
Otro blanco que es fosforilado
es el canal de la Ca2+ tipo L, por acción de las
catecolaminas. Favorece al cargar con más Ca2+ en el
retículo y puede modificar la pendiente de repolarización.
Características de los canales de calcio tipo L y tipo T
- Tipo L:
- Umbral de activación de -30 mV.
- Umbral de cierre de -40 mV.
- Inactivación lenta, dependiente de voltaje.
- Dependiente de Ca2+.
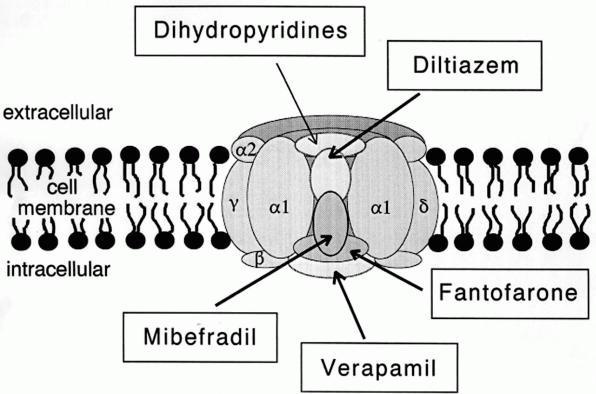 |
| Canal de calcio tipo L (dominio público) |
- Tipo T:
- Umbral de activación de -60 mV.
- Umbral de cierre de -90 mV.
- Inactivación rápida dependiente de voltaje.
- Dependiente de tiempo.
Influencias autonómicas sobre la VRC:
 |
| Inervación autonómica del corazón (OpenStax College, Anatomy and Phisiology, 2013. CC BY 3.0). |
Las principales áreas de
inervación simpática y parasimática en el corazón en el techo del
atrio derecho y hacia el piso en el septum del atrio derecho, donde
existe el sistema de marcapasos y el sistema excito-conductor.
Las fibras simpáticas van por
la región epicárdica y las parasimáticas por la endocárdica e
ingresan también hacia al miocardio contráctil, pero la densidad de
la sinapsis es 1.000 veces menor en el tejido contractil versus el
que lleva el ritmo.
La base molecular para la
modificación del ritmo cardíaco son os potenciales espontáneos de
las células marcapasos, de los cuales es responsable el canal funny
(canal catiónico inespecífico que permite el paso de iones Na+
y K+).
La actividad de este canal aumenta en la medida que el potencial es
más negativo (la noradrenalina y la acetilcolina también tienen
efecto sobre esta actividad). Este canal tiene dependencias de AMPc.
Mecanismo de acción de las catecolaminas:
Actúan a través de receptores
beta-adrenérgicos presentes en las células marcapasos presentes en
las células contráctiles.
Estos receptores
beta-adrenérgicos están acoplados a proteínas G que actúan
aumentando la concentración de AMPc.
Este mecanismo se puede
manipular farmacológicamente sin modificar el tono.
Mecanismo de acción de la acetilcolina:
El sistema parasimpático actúa
a través de receptores muscarínicos tipo M2 que tiene acopladas una
proteína G inhibitoria.
La
ACh disminuye la actividad de los canales funny,
o también disminuye o cambia su dependencia de voltaje.
 |
| Receptor muscarínico M2 (A2-33, Wikimedia Commons, CC BY-SA 3.0) |
La pendiente del potencial
marcapasos:
- Aumenta en presencia de catecolaminas.
- Disminuye en presencia de acetilcolina.
Modificando la frecuencia de
producción de potenciales marcapasos.
Incx
provoca una corriente de Ca2+
a través de un intercambiador Na+/Ca2+,
que se ve aumentado en pacientes con insuficiencia cardíaca. Tiene
su máxima actividad durante la repolarización (importante, por
ejemplo, en un paciente que tenga reducción de la actividad de su
bomba de ión calcio del retículo).
El potencial diastólico máximo
puede variar por el sistema nervioso parasimático. Cuando este
potencial es negativo, a una misma pendiente del potencial marcapasos
le va a tomar más tiempo alcanzar el potencial umbral y, por lo
tanto, va a reducir la frecuencia de los potenciales espontáneos.
El
receptor mucarínico no siempre está acoplado a una proteína G
inhibitoria, sino que atambién a otra proteína GK, propia del
miocardio en los nodos SA y AV. La proteína GK está acoplada a un
canal de K+,
IK-ACh,
que provica una corriente de K+,
hiperpolarizando a la célula o “haciendo más negativo el
potencial diastólico máximo.”
Relación entre el potencial de acción miocárdico y el electrocardiograma (ECG):
Cuando se ponen electrodos en
la superficie de la piel, resulta que aparece una serie de
deflexiones características que son producto de la forma secuencial,
ordenada, y del número de células que forman potenciales de acción
al mismo tiempo, que es lo que se muestra en el electrocardiograma.
 |
| Paciente con un monitor de Holter (un aparato electrocardiográfico portátil) (Usuario Jason7825, Wikimedia Commons. CC-BY-SA-3.0,2.5,2.0,1.0) |
No hay comentarios:
Publicar un comentario